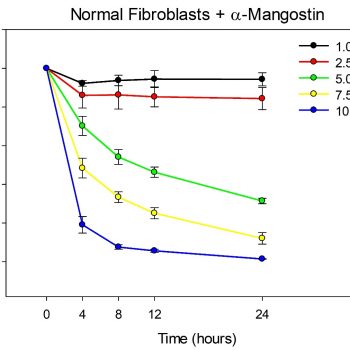
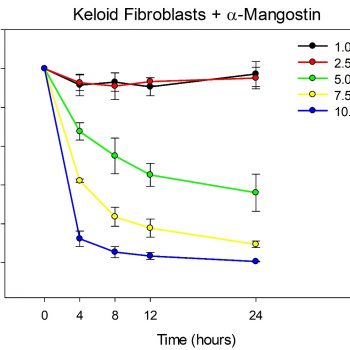
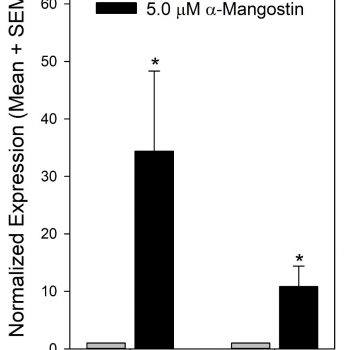
VISUAL ABSTRACT
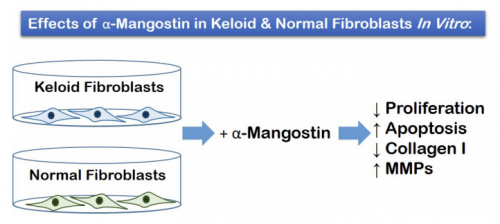
ABSTRACT
Keloids are disfiguring lesions that result from an abnormal wound healing process. Despite the availability of numerous therapeutic options, keloids remain challenging to treat and often recur after therapy. α-Mangostin, a natural xanthone isolated from the fruit of the Mangosteen tree, has been used for centuries in many Southeast Asian nations for medicinal purposes, and has gained attention more recently due to its anti-inflammatory, antimicrobial, and antioxidant properties, with numerous studies suggesting possible anticarcinogenic activities. Hypothetically, α-mangostin may have therapeutic value for keloid suppression. To investigate this hypothesis, the effects of α-mangostin on fibroblast proliferation, apoptosis, and gene expression in vitro were analyzed. Dermal fibroblasts were isolated and cultured from normal human skin and excised keloid lesions (3 donors each), and were treated with multiple doses of α-mangostin in vitro. Dose-dependent decreases in proliferation in keloid and normal fibroblasts were observed following treatment with α-mangostin. The α-mangostin treated fibroblasts displayed significantly increased expression of C/EBP homologous protein (CHOP), which mediates endoplasmic reticulum stress-induced apoptosis, suggesting increased apoptosis. In addition, numerous changes in gene expression were observed in α-mangostin-treated keloid fibroblasts, including decreased expression of collagen type I alpha 1 chain (COL1A1) and increased expression of matrix metalloproteinase 1 (MMP1), MMP3, and MMP13. Secretion of pro-collagen I was decreased, and secretion of MMP1 and MMP3 were increased, in α-mangostintreated fibroblasts. The results suggest that α-mangostin may exhibit antiproliferative, proapoptotic, and antifibrotic activities in keloid and normal fibroblasts. Thus, this study suggests that further investigation is warranted to fully explore the therapeutic potential of α-mangostin for suppression of keloid development.
Introduction
Keloids are fibroproliferative lesions that can result from an abnormal wound healing response. Although historically considered an extreme form of pathological scarring, the term “keloid disorder” has been proposed to distinguish these lesions from other types of scars, such as hypertrophic scars.1,2 Keloids are characterized by excessive and abnormal deposition of collagen, with an elevated ratio of type I to type III collagen.3 Unlike hypertrophic scars, which are raised but limited to the original wound area, keloids tend to bulge over the original wound margin and can continue to increase in size indefinitely.4 Keloids are relatively common, although the incidence varies among different populations, with increased risk of keloid formation in individuals of African, Asian, and Hispanic descent compared with those of European ancestry.1,2,5 Keloids are disfiguring and can cause pain, itching, decreased range of motion, and impaired psychosocial well-being.6-9 Keloids are extremely resistant to treatment, usually recurring even after surgical excision.1 There are many different treatment modalities for keloids, but most have limited long-term success, and highquality evidence supporting clinical efficacy is lacking for most therapies.1,10,11 Improved therapeutic interventions are needed to suppress keloid pathology and alleviate patient morbidity.
The molecular mechanisms leading to keloid formation remain incompletely understood, despite numerous studies reporting gene expression abnormalities in keloids and keloid-derived cells.12-15 The fact that keloids have an increased prevalence in certain populations, and tend to run in families, suggests a genetic predisposition.16 Although numerous chromosomal loci have been associated with keloid risk, no causative gene has been identified to date, and it is likely that keloid susceptibility is a multigenic trait.16,17 Keloid fibroblasts exhibit increased proliferation, decreased apoptosis, increased expression of type I collagen, and exaggerated responses to transforming growth factor beta 1 (TGF-β1) signaling.3,18-20 In addition, keloid fibroblasts exhibit increased expression of proinflammatory cytokines and altered regulation of genes in multiple signaling pathways, including TGF-β1 and WNT/β-catenin pathways. 13-15,21 Agents that reverse this fibrotic phenotype are expected to have potential therapeutic value for keloid suppression. There has been increasing interest in recent decades in the medicinal uses of natural, botanical dietary supplements, due in part to their perceived healthpromoting qualities.22 Many botanical supplements have demonstrated medicinal properties, such as anti-inflammatory, anti-infective, and anti-oxidant properties, and there has been a great deal of research into potential therapeutic applications of phytochemicals. One plant that has a long history of medicinal use is Garcinia mangostana, also known as mangosteen, a tropical tree native to Southeast Asia. The fruit of the mangosteen tree has been used for centuries as a medicinal agent, particularly for treatment of wounds and skin infections, as well as abdominal pain, diarrhea, dysentery, and ulcers.23,24 The pericarp of the fruit is rich in xanthones, a class of polyphenolic compounds with numerous biological activities; among these, α-mangostin is a major constituent.24 Because α-mangostin has been reported to exhibit numerous pharmacological properties, it has been explored as an anti-cancer agent. Evidence from in vitro and preclinical in vivo studies suggests that α-mangostin has anticarcinogenic activities affecting tumor initiation, promotion, and progression.25 α-Mangostin has potent antioxidant activities, which include scavenging of free radicals and modulation of oxidative stress-related enzymes.23,25 In addition, α-mangostin has antiproliferative and proapoptotic activities.25 For example, α-mangostin reduced proliferation via cell cycle arrest in a human colon cancer cell line, in part through downregulation of cyclin protein levels, and induced apoptosis in human leukemia cell lines and a human osteosarcoma cell line.26- 29 Anti-inflammatory activity of α-mangostin has been demonstrated in multiple human cell lines from different tissues; for example, α-mangostin reduced expression of the proinflammatory mediators tumor necrosis factor alpha (TNFα) and interleukin 8 (IL8) in HepG2 hepatocyte carcinoma cells, Caco-2 colorectal adenocarcinoma cells, and HT-29 colon cancer cells.30 Antifibrotic activities have been demonstrated in hepatic stellate cells in vitro, suggesting therapeutic potential for suppression of liver fibrosis.31,32 For example, treatment of LX2 hepatic stellate cells with α-mangostin decreased proliferation, TGF-β1 secretion, and decreased expression of collagen type I alpha 1 chain (COL1A1), alpha smooth muscle actin, and phosphorylated SMAD family member 3 (SMAD3), considered markers of fibrogenesis.31,32
Because of the antiproliferative, anti-inflammatory, proapoptotic, and anti-fibrotic activities of α-mangostin that were previously described, it may represent an attractive new therapeutic for suppression of keloid scarring. To begin to explore this possibility, we investigated the effects of α-mangostin in keloid-derived cells in vitro. Fibroblasts cultured from keloid lesions and normal human skin were treated with α-mangostin; proliferation was quantified, and expression levels of fibrotic markers and other genes implicated in keloid pathology were analyzed.